
Willandra Lakes Hominid 50 was recovered from a deflating land surface in the Garnpung/Leaghur Lake region of south-western New South Wales, with the first published report in Flood (1983). This skeleton has not been reliably dated, has not been formally described, and is probably pathological. These circumstances result in some unease over the extreme claims made about the relevance of WLH 50 to interpretations of the Australasian evolutionary sequence (Stringer 1992; Brown 1992; Stringer and Bräuer 1994). In particular WLH 50 regularly appears as a corner stone in arguments for evolutionary continuity between the Indonesian and Australian regions published over the last two decades (Thorne 1984; Wolpoff 1992, 1995; Thorne and Wolpoff 1992; Frayer et al. 1993; Frayer et al. 1994; Hawks et al. 2000) which is an unusual circumstance for an undescribed and poorly provenanced fossil.
Attempts to date WLH 50 have obtained controversial results. Initial attempts to obtain a radiocarbon date achieved a result much younger than expected. It is possible that the specimen was contaminated and material other than collagen was dated. It is also possible that the fossil is a lot younger than some people would like. More recently Caddie et al. (1987) reported an electron spin resonance date on bone of 29,000 ±5000 years BP for WLH 50. Grün and Schwartz (1987) argue that while ESR can produce reliable dates on dental enamel bone is not a suitable material for ESR. Surprisingly, in diagrams, or tables, where WLH 50 is listed along with dated terminal Pleistocene Australian skeletons Wolpoff and Thorne (Thorne and Wolpoff 1992; Wolpoff 1995) either describe WLH 50 as the most archaic or position it as the earliest human fossil from Australia. There is no published evidence to support this line of argument. Simpson and Grün (1998) have published a U-series date of 12 to 18 ka for Wilandra Lakes 50, and recently Grünet al. (2011) conclude that "Gamma spectrometry was carried out on the WLH 50 calvaria and TIMS U-series analysis on a small post-cranial bone fragment. OSL dating was applied to a sample from Unit 3 at a level from which the WLH 50 remains may have eroded, as well as from the underlying sediments. Considering the geochemistry of the samples analysed, as well as the possibility of reworking or burial from younger layers, the age of the WLH 50 remains lies between 12.2 ± 1.8 and 32.8 ± 4.6 ka (2-σ errors)".
![]()




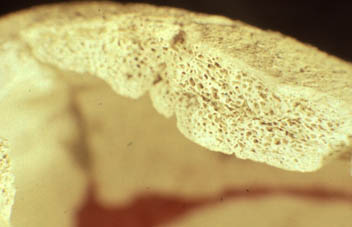
(copyright Peter Brown 1982)
![]()
WLH 50 consists of a fragmentary cranial vault, with damage to the basal and temporal segments, some facial fragments, parts of an elbow joint and some smaller postcranial pieces. The most striking feature of the cranial vault, malar fragments and elbow is of great size. Although glabella is not preserved, maximum cranial length can be estimated (±3 mm) to 212 mm, with a maximum cranial breadth of 151 mm and maximum supraorbital breadth greater than 131 mm. These dimensions exceed the recorded Aboriginal range of variation (Brown 1989). Even with the pathologically thickened vault, discussed below, endocranial volume was approximately 1540 ml compared with the Holocene Aboriginal male mean of 1271 ml Brown (1992b) . The extremely large size of WLH 50 should be of some concern to those who argue that this skeleton is in some manner representative of 'Late Pleistocene' Australians (Thorne 1984; Wolpoff 1992, 1995; Thorne and Wolpoff 1992; Frayer et al. 1993; Frayer et al. 1994).
The cranial vault of WLH 50 is long, broad and high. The frontal bone, continuous with the supercilliary ridges, is flattened in its anterior two thirds but angles sharply upwards as it moves away from the supraorbital region. The posterior third curves towards bregma and the frontal bone does not form a low continuous arch as in middle Pleistocene Indonesian hominids. There is no supraorbital torus, the supercilliary ridges are small for an Aboriginal male cranium of this size, and the zygomatic trigones are poorly developed. There is certainly no backward extension of the supraorbital region at the outer corner of the orbit, along the temporal line, forming a knob-like trigone as is evident in the Ngandong and Sangiran hominids. A cruciform eminence is not present at bregma.
Sagittal curvature of the parietal bones (curvature index 19.2) is close to the Holocene Aboriginal male mean of 19.8, however maximum bi-parietal breadth is located in a much more inferior position than is common in terminal Pleistocene or Holocene Aboriginal crania. What is preserved of occipital morphology falls within the range of normal Aboriginal male variation, apart from overall size. The occipital torus, although somewhat eroded, clearly did not correspond either in size or morphology with the same region in the Ngandong or Sangiran hominids. Although the inion region was undoubtedly large, what is preserved does not resemble the pronounced inferiorly pointing triangle present at Ngandong. Similarly the occiput does not bulge along the lambdoid suture as it does at Ngandong. The morphology of the supramastoid crest and sulcus and mastoid crest are consistent with those in recent Aboriginal male crania and distinct from Ngandong. The zygomatic fragments are large, but of completely modern human morphology.
| bregma-lambda | 130 | max cranial br. | 151 |
| parietal subt. height | 25 | bi-asterion | 127 |
| bregma-subtense | 61 | glabella-opisthocranion | (212) |
| bregma-asterion | 151 | thickness mid-frontal | 16.5 |
| lambda-asterion | 81 | thickness bregma | 17.2 |
| lambda-inion | (74) | thickness at vertex | 13.9 |
| bregma-inion | (168) | thickness at parietal boss | 14 |
| asterion-inion | (71) | thickness at lambda | 16.2 |
| max supraorbital | (131) | thickness at asterion | 16 |
| min post orbital | 109 | thick. at int. occipital prot. | (21) |
| max bi-parietal | 139 |
Perhaps the most unusual aspect of WLH 50 is the manner in which it has been used to support arguments for evolutionary continuity between middle Pleistocene Javan hominids and terminal Pleistocene Australians. In the first detailed account of their regional continuity model Thorne and Wolpoff (1981) assume that 'terminal Pleistocene' Australians were a fairly homogenous group and are suitably represented by Kow Swamp and the undated and poorly provenanced Cohuna cranium. As I have discussed previously (Brown 1992a) well dated terminal Pleistocene crania from Lake Mungo (26,000 years BP) and Keilor (12,000 years BP) were excluded from Thorne and Wolpoff's comparison. In retrospect this appears to be because the morphology of these crania did little to support their argument. The same applies to the use of WLH 50. If their regional comparison has substance surely it should include the earliest securely dated Australian fossils. As it happens it does not. Comparisons should be between the Sangiran and Ngandong hominids and Lake Mungo 1 and Keilor. Comparative photographs are included below. Draw your own conclusions. The photographs below are all to the same scale.
![]()



![]()
Brown, P. 1989. Coobool Creek. Terra Australis 13. Department of Prehistory, Australian National University.
Brown, P. 1992a. Recent human evolution in East Asia and Australasia. Transactions of the Royal Society London Series B 337: 235-242.
Brown, P. 1992b. Post-Pleistocene change in Australian Aboriginal tooth size: dental reduction or relative expansion? In T. Brown and S. Molnar (eds) Craniofacial variation in Pacific population, pp. 33-52. Anthropology & Genetics Laboratory, Department of Dentistry, University of Adelaide.
Brown, P. 1994. Cranial vault thickness in Asian Homo erectus and Homo sapiens. Courier Forschungsinstitut Senckenberg 171: 33-46.
Caddie, D., Hunter, D., Pomery, P. and Hall, H. The ageing chemist - can electron spin resonance (ESR) help? In W. Ambrose and J. Mummery (eds.) Archaeometry: further Australasian studies, pp. 156-166. Department of Prehistory, Australian National University.
Frayer, D., Wolpoff, M., Smith, F., Thorne, A. and Pope, G. 1993. The fossil evidence for modern human origins. American Anthropologist 95: 14-50.
Frayer, D., Wolpoff, M., Thorne, A., Smith, F. and Pope, G. 1994. Getting it straight. American Anthropologist 96: 424-430.
Grün, R. and Schwartz, H. 1987. Some remarks on 'ESR dating of bones'. Ancient TL 5: 1-9.
Grün, R. et al., 2011. Stratigraphy and chronology of the WLH 50 human remains, Willandra Lakes World Heritage Area,Australia. Journal of Human Evolution 60: 597-604.
Hawks, J. et al., 2000. An Australasian test of the recent African origin theory using the WLH-50 calvarium. Journal of Human Evolution 39: 1-22.
Simpson, J.J. and Grün, R. 1998. Non-destructive gamma spectrometric U-series dating. Quaternary Science Reviews (Quaternary Geochronology) 17 (11).
Stringer, C.B. 1992. Replacement, continuity and the origin of Homo sapiens. In G. Bräuer and F. Smith (eds) Continuity or replacement, pp.9-24. A.A. Balkema.
Stringer, C. and Bräuer, G. 1994. Methods, missreading and bias. American Anthropologist 96: 416-424.
Thorne, A. and Wolpoff, M.H. 1981. Regional continuity in Australasian Pleistocene hominid evolution. American Journal of Physical Anthropology 55: 337-349.
Thorne, A. and Wolpoff, M.H. 1992. The multiregional evolution of humans. Scientific American 266: 76-83.
Webb, S. 1990. Cranial thickening in an Australian hominid as a possible palaeoepidemiological indicator. American Journal of Physical Anthropology 82: 403-412.
Wolpoff, M.H. 1992. Theories of modern human origins. In G. Bräuer and F. Smith (eds) Continuity or replacement, pp.25-64. A.A. Balkema.
Wolpoff, M.H. 1995. Human Evolution 1996 edition. McGraw Hill.
![]()