![]()

![]()
In August 2003, excavations directed by the late Michael Morwood (UNE) and the late Radien Sajono (Arkeologi Nasional, Jakarta), in the Pleistocene deposits at Liang Bua Cave, western Flores, uncovered the partial skeletal remains of a small-bodied and small-brained hominin, as well as a large number of stone artefacts and the bones of extinct fauna, including Stegodon (elephant relative) (Morwood et al., 2004). The combination of unique, derived and primitive traits in the LB1 skeleton resulted in its allocation to a new species, Homo floresiensis (Brown et al., 2004). In the following year, these discoveries were further enhanced with the excavation of additional skeletal components from as many as nine individuals (Morwood et al., 2005), and most of the hominin fossils, as well as their geochronological and cultural contexts, have now been published in detail (Brown et al, 2004; Falk et al., 2005, 2009; Tocheri et al., 2007; Brown and Maeda, 2009; Larson et al., 2009; Moore et al., 2009; Morwood and Jungers, 2009; Roberts et al., 2009; Jungers et al., 2009; Westaway et al., 2007; Brown 2012).
Currently (2012), the evidence for H. floresiensis at Liang Bua, including the associated cultural deposits, is dated to between 95,000 and 17,000 years ago. However, in common with most caves, the depositional history at Liang Bua is complex and there were periods when the cave was flooded and sections of the deposit eroded and redeposited. Future excavation and analysis of stratigraphic relationships may result in some adjustment of the dates of the Pleistocene deposits, and the evidence for the apparently simultaneous extinction of H. floresiensis and Stegodon. It is unlikely that the evidence for extinction will draw closer to the Holocene, but a substantial increase in age would not be a great surprise.
Moving ahead nearly two decades after the original publication, more detailed archaeological and geochronological research at Liang Bua have modified the initial conclusions about the age of the Homo floresiensis and Stegodon deposits in the cave and the potential contemporaneity with modern humans (Sutikna et al., 2018; Sutikna et al., 2016). The revised stratigraphic and chronological evidence no longer supports the initially inferred ages for the H. floresiensis holotype (LB1), ~18 thousand calibrated radiocarbon years before present (kyr cal. BP), or the time of last appearance of this species (about 17 or 13-11 kyr cal. BP). Instead, the skeletal remains of H. floresiensis and the deposits containing them are dated to between about 100 and 60 kyr ago, with the stone artefacts attributable to this species ranging in age from about 190 to 50 kyr (Sutikna et al., 2016). The simple explanation for why the initial dates from Liang Bua were revised is that a lot more was known about the stratigraphy of the cave deposits in 2015 than in 2003. Cave deposits are usually stratigraphically complex, with flooding, erosion, reworking and redeposition of different parts of the cave floor is common. Objects may move both vertically and horizontally through a deposit due to these processes, as well as treadage, animal burrowing and gravity. These are all issues at Liang Bua. The most reliable guide to whether faunal remains are in the original, undisturbed stratigraphic position, is when skeletal elements are still articulated, or in proximity. This was the situation with the LB1 H. floresiensis skeleton. It is also likely that some of the initial enthusiasm to add human-like cultural baggage, including the use of fire and stone tool cut marks on Stegodon bones, may have been an overreach. Some of the archaeologists (Mike Morwood) and geochronologists (Richard Roberts) believed that the Liang Bua hominins would have greater impact if they were firmly intertwined with the human story, rather than just being recently extinct, tool-making apes. Natural processes can produce scratches on a bones surface, as can the teeth of carnivores and rodents. A detailed analysis of the evidence for the use of fire by H. floresiensis has not been published (2022) and the "cut marks" that I have seen on Stegodon bones (2003-2005) could have resulted from processes other than hominin use of stone tools.
![]()
 |
 |
 |
|---|---|---|
3D reconstruction of the LB1 H. floresiensis cranium and mandible based on low resolution CT data (1 mm slice interval). Rotation on the horizontal axis. |
3D reconstruction of the LB1 H. floresiensis cranium and mandible based on low resolution CT data (1 mm slice interval). Rotation on the vertical axis. | Cranium and mandible of LB1 H. floresiensis after initial cleaning and reconstruction in 2003. |
(copyright Peter Brown 2003-2012)
![]()
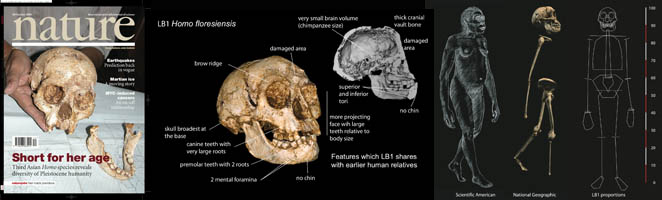
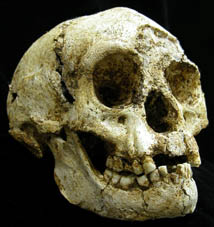
The most complete skeleton (LB1) comprises a cranium and mandible, with bones of the right arm, pelvis, legs and feet, as well as less well preserved parts of the rest of the skeleton (Brown et al. 2004; Morwood et al. 2005; Larson et al. 2009; Jungers et al. 2009). Descriptive comparisons of the cranium, mandible and teeth have highlighted the unique combination of anatomical characteristics. For example, the brain volume (380-420 cubic centimeters) is similar to early australopithecines and living chimpanzees, the mandibular first premolars are elongated, asymmetrical and have complex roots (common in australopithecines and early Homo), and the mandible also has internal buttressing (reinforcement) most similar to early australopithecines (Brown et al. 2004; Brown and Maeda 2009; Brown 2012).
Although only one cranium (LB1) has so far been found (2012) there is postcranial and dental evidence from multiple individuals. For instance, there are three left mandibular premolars with the same distinctive crown and root anatomy (Brown and Maeda 2009; Brown 2012) and two mandibles (LB1 and LB6) that share a similar small size and set of primitive anatomical characteristics (Brown and Maeda 2009). The postcranial skeletal elements are all from extremely short hominins, with the estimated stature for LB1 of 105cm making it the “basketball player” of the group. Features shared with australopiths include relatively long arms, flared pelvis, relatively short legs and the shape of some bones of the wrist. The latter possibly limiting their manipulative skills. This contrast with the feet of LB1 that are similar in overall form to those in H. sapiens (big toe aligned with the others and foot clearly adapted for terrestrial bipedal walking), although relatively long as in chimpanzees and australopiths. Research by William Jungers and colleagues (Jungers et al., 2009) also suggests that the foot of LB1 lacks the longitudinal arch found in H. erectus and modern humans.
![]()
 |
 |
|---|---|
 |
(copyright Peter Brown 2004-2005)
![]()
However, the allocation of these fossils to a new species has not gone unchallenged, with several independent teams arguing that morphologically and metrically the LB1 skeleton falls within the range of modern humans with particular developmental disorders or syndromes (Weber et al., 2005; Jacob et al., 2006; Martin et al., 2006; Richards, 2006; Hershkovitz et al., 2007, 2008; Vannuccia et al., 2011). In turn, these claims for pathology have been reviewed and found to be insubstantial and speculative, with the critics unable to present a modern human skeleton with the attributes of H. floresiensis (Falk et al., 2007, 2009, 2010; Brown and Maeda, 2009; Jungers et al., 2009; Aiello, 2010; Kaifu et al., 2010; Baab and McNulty 2009; McNulty and Baab, 2010; Brown 2012; Baab et al 2016).

(copyright Peter Brown 2005)
Stated concerns for those favouring a pathological interpretation of H. floresiensis include the association between the cultural remains and hominin fossils, the intellectual capabilities of a hominin with a brain the size of a chimpanzee, and a belief that Homo sapiens may have been contemporaneous with LB1 on Flores. The latter presumably driven by the archaeological evidence for modern humans on the Sahul Shelf and Bismark Archipelago by 45-46 ka (thousands of years ago) (O’Connell et al., 2010) and on Timor at >42 cal BP (calibrated years before present) (O’Connor 2007). These claims were recently reviewed by Brown (2012), who concluded that there is currently no archaeological or human skeletal evidence from Flores or Liang Bua, which indicates H. sapiens was on the island during the Late Pleistocene.
In the original description and interpretation of Homo floresiensis (Brown et al., 2004) it was concluded that this small-bodied and small-brained hominin was either the result of insular dwarfing of an Homo erectus ancestor on Flores, or was the descendant of an australopith/early Homo-like ancestor, with limited evolutionary change on Flores. With the description of additional H. floresiensis fossils in 2005 (Morwood et al., 2005), including the right arm, Peter Brown concluded that skeletal dimensions, scaling trajectories and the combination of plesiomorphic and unique anatomical traits, were not consistent with dwarfing in H. erectus. This theme was further examined in Brown and Maeda (2009). Resolution of this issue will depend upon the discovery of mid-Pleistocene hominin skeletal materials on Flores, or adjacent islands.
The conclusion that the first hominins to reach Flores were already small-bodied, and not the result of in situ island dwarfing from large-bodied H. erectus ancestors, may be supported by more recent discoveries from the middle Pleistocene site of Mate Menge, Ola Bula Formation in the So'a Basin (Kaifu et al. 2024). Fragments of a mandible and humerus from small-bodied hominins have been argued to be both similar in size and anatomy to H. floresiensis from Liang Bua. Kaifu and colleagues chosen interpretation is that the Mate Menge fossils indicate early and rapid dwarfing of H. erectus on Flores, providing the ancestors of H. floresiensis. This interpretation runs contrary to the substantial English language literature, for instance Brown and Maeda (2009), Argue, et al. (2017) phylogenetic analysis of cranial and postcranial traits, and articles on the postcranial skeleton of LB1 in the Journal of Human Evolution volume 57 number 5 special edition on Liang Bua, indicating that the most likely ancestor of H. floresiensis was small-bodied and small-brained early Homo. The combination of plesiomorphic and unique traits present in H. floresiensis are not reproduced in known examples of Homo erectus, or the fragments from Mate Menge, and there is currently no evidence of a large-bodied, middle-Pleistocene ancestor, H. erectus or anything else, on Flores. An additional concern is the age and faunal associations of the Mate Menge hominin fragments. The bones in the site are the result of secondary deposition, where bones have been exposed on the surface, washed and tumbled into a gully, broken and not articulated (Powley et al. 2021). The remains of Stegodon and stone tools have been reported from this site (Morwood, et al. 1998), but it is possible that the hominin fossils are not contemporary with them. If the hominin fossils are indeed dated to ~700,000 years, and from a hominin of similar body size to LB1 H. floresiensis, then surely given the existing literature, the most parsimonious explanation is that the first hominins to reach Flores were small-bodied, small-brained, and with a host of cranial, postcranial and dental traits shared with African early Homo and Ausralopithecines. While I suggested that insular island dwarfing was one possible explanation for the anatomical characteristics found in LB1 H. floresiensis (Brown et al. 2004) I no longer think that this is a realistic option.
 |
 |
|---|---|
The only subadult evidence for H. floresiensis at Liang Bua are for the extremely small LB4 tibia and radius. Note the relatively long radius. |
Mandibular and dental remains of pygmy Stegodon from Liang Bua. |
(copyright Peter Brown 2005)
 |
|---|
 |
|---|
![]()
 |
craniometric data for LB1 H. floresiensis, H. erectus and H. habilis |
|---|
![]()
References
Aiello LC. 2010. Five years of Homo floresiensis. Am J Phys Anthropol 142(2):167-179.
Baab, K.L., Brown, P., Falk, D.j, Richtsmeier, J.T., Hildebolt, Smith, K., and Jungers, W. 2016. A Critical Evaluation of the Down Syndrome Diagnosis for LB1, Type Specimen of Homo floresiensis.PLOS ONE https://doi.org/10.1371/journal.pone.0155731
Martin RD, Maclarnon AM, Phillips JL, and Dobyns WB. 2006a. Flores hominid: new species or microcephalic dwarf? The Anatomical Record 288A(11):1123-1145.
Martin RDR, Maclarnon AMA, Phillips JLJ, Dussubieux LL, Williams PRP, and Dobyns WBW. 2006b. Comment on "The Brain of LB1, Homo floresiensis". Science (New York, NY) 312(5776):999.
McNulty, K.P., Baab, K.L., 2010. Keeping asymmetry in perspective: A reply to Eckhardt and Henneberg. Am. J. Phys. Anthropol. 143, 337-339.
O'Connell, J.F., Allen, J., Hawkes, K., 2010. Pleistocene Sahul and the origins of seafaring, in: Anderson, A., Barrett, J., Boyle, K. (Eds.), The global origins and development of seafaring. McDonald Institute for Archaeological Research,, Cambridge, pp. 57-68.
O'Connor S. 2006. New evidence from East Timor contributes to our understanding of earliest modern human colonisation east of the Sunda Shelf. Antiquity 81:523-535.
Sutikna, T., Tocheri, M.W., Faith, J.T., Awe, R.D., Meijer, H.J., Saptomo, E.W., Roberts, R.G., 2018. The spatio-temporal distribution of archaeological and faunal finds at Liang Bua (Flores, Indonesia) in light of the revised chronology for Homo floresiensis. J Hum Evol 124, 52-74.
Sutikna, T., Tocheri, M.W., Morwood, M.J., Saptomo, E.W., Awe, R.D., Wasisto, S., Westaway, K.E., Aubert, M., Li, B., Zhao, J.-x., Storey, M., Alloway, B.V., Morley, M.W., Meijer, H.J., van den Bergh, G.D., Grun, R., Dosseto, A., Brumm, A., Jungers, W.L., Roberts, R.G., 2016. Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature 532, 366-369.
Tocheri MW, Orr CM, Larson SG, Sutikna T, Jatmiko, Saptomo EW, Due RA, Djubiantono T, Morwood MJ, and Jungers WL. 2007c. The Primitive Wrist of Homo floresiensis and Its Implications for Hominin Evolution. Science. p 1743-1745.