![]()



![]()
The Lake Mungo 3 skeleton was discovered by Jim Bowler. In February 1974, after prolonged rain in 1973, Bowler noticed the exposed left side of a carbonate encrusted human cranium 500m east of the Mungo 1 cremation site. Later that month the extended burial that was Lake Mungo 3 was excavated by a team from the Australian National University (Bowler and Thorne 1976). When first published this burial had not been directly dated but Bowler and Thorne argued on the basis of geomorphological criteria and stratigraphic association with Mungo 1 that an age of 28,000 to 32,000 years BP was probable (1976: 136-138). In 1987 Caddie et al. reported an electron spin resonance date on bone of 31,000 ±7000 years BP for Lake Mungo 3. Grün and Schwartz (1987) argue that while ESR can produce reliable dates on dental enamel bone is not a suitable material for ESR. Most recently Thorne et al. (1999) report that the age of the Lake Mungo 3 burial, as determined by the combined results of electron spin resonance (ESR) on dental enamel, U-series on calcite crust covering the skeleton and OSL on the sediment surrounding the skeleton indicate colonisation during or before oxygen isotope stage 4 (57,000-71,000). These dates make little sense given the original stratigraphic argument and the absence of archaeological materials of similar age from the Willandra Lakes region. The bottom of the Mungo Unit from which LM3 was recovered has been securely dated to 43,000 years. Given that the grave of LM3 must have been dug from a higher, and therefore younger, land surface there is little possibility that the LM3 burial can be older than 43,000 years. Critiques of Thorne et al. (1999) dating of LM3 by Gillespie and Roberts (2000), and Bowler and Magee (2000), have been published in the Journal of Human Evolution. In the same volume Brown )examines Thorne et al. (1999) claims for morphological gracility and male sex for LM3. There is a repy to the two dating critiques by Grün et al. (2000). A review of the debate can be found in Brown (2000b), with further contributions by Thorne and Curnoe (2000) and Brown and Gillespie(2000). Bowler et al. (2003) argue that the OSL samples used to date LM3 by Thorne et al (1999) had little, if any, stratigraphic relationship with the original burial. They report the results of 25 new OSL dates which suggest LM3 can not be older than 50,000 years BP. While the dating of the Lake Mungo stratigraphic sequence now seems more secure I don't believe that Bowler et al (2003) have established a minimum age for the burial, only a possible maximum. When LM3 was discovered it had been exposed by erosion. You can only speculate at the nature of the original deposit above the burial, and the land surface from which it was intered. I suspect that LM3 was buried during a period in which the nearby lake was full of water, with the environment most suited for human habitation, but lake levels fluctuated over a a long period of time (Bowler et al. 2003). A range of 15,000 to 50,000 appears possible (Gillespie 2002).
Most recently LM3 has attracted attention due to the claim by Gregory Adcock and co workers that they had extracted mtDNA from fragments of LM3's skeleton (Adcock et al. 2001). The authors identify differences between the mtDNA of LM3 and mtDNA of living Aborigines and terminal Pleistocene skeletons from Kow Swamp. They conclude that the mtDNA of LM3 belonged to a lineage that only survives as a segment inserted into chromosome 11 of the nuclear genome which is now widespread among human populations. The LM3 lineage probably diverging before the most recent common ancestor of contemporary human mitochondrial genomes. For Adcock et al. (2001) this implies that the deepest known mtDNA lineage from an anatomically modern human occurred in Australia.
Their have been some criticisms of their claims, largely as their results have not been independently tested, and contamination has proved to be an enormous problem in fossil DNA research (Nolch 2001; Cooper et al. 2001). Research in other parts of the world suggests that the hot, but variable, climate of the Willandra Lakes is unlikely to be conducive to organic preservation, particularly at the date of 60 kyr claimed by Adcock et al (2001). In Europe and north Africa, for instance, DNA dated to more than 10 kyr has only been recovered from cold environments, with climates similar to the Willandra Lakes not yielding genetic material. There may, of course, be something unusual about the preservation of LM3, or the great care in which it was processed by Greg Adcock, but at present the preservation of mtDNA in LM3 is at odds with what is known from the rest of the world. What is currently known about aDNA survival and post-depositional thermal history of the site make it extremely unlikely that aDNA was amplified from skeletal material at either Lake Mungo, or Kow Swamp (Smith et al. 2003).
Other criticisms of Adcock et al. (2001) have focused on their selective use of the literature and the way in which they have interpreted their data. For instance the authors state that LM3 is approximately 60 kyr old and ignore the literature arguing that it must be younger than 40 kyr (Bowler and Magee 2000; Gillespie and Roberts 2000; Brown 2000b; Brown and Gillespie 2000), they indicate the LM3 is morphologically gracile compared with other Pleistocene Australians when the only tests of this statement have demonstrated that it is not true(Brown 1987; Brown 2000a), and also state that most workers have linked variation in Aboriginal morphology to migration by morphologically distinct groups. A review of the historical literature demonstrates that this interpretation is incorrect (Thorne 1976; Brown 1997). Cooper et al (2001) argue that standard DNA authentication criteria, including independent replication, biochemical studies of bone preservation, and cloning of DNA sequences should have been used to rule out possible contamination from modern handling. They all also suggest that the high proportion of cytosine-thymidine transitions between LM3 and the reference sequence correspond well with the cytosine deamination common in damaged DNA. Adcock et al. (2001b) have responded to some of these criticisms. Recently, Thomas et al (2003) have examined the distribution patterns of postmortem damage in human mitochondrial DNA. They suggest that postmortem damage may explain many unusual results obtained from ancient human remains when appropriate techniques were not followed, including the "Mungo man" sequences published by Adcock et al. (2001).
In the long run uncertainty over the role of contamination in the reported mtDNA from LM3 and Kow Swamp will only be clarified through independent testing. Unfortunately, this may prove impossible as the Kow Swamp skeletons were all supposed to have been returned for reburial in 1986, and it is unlikely that Aboriginal communities will be sympathetic to further testing of the LM3 skeleton.
![]()
 |
 |
 |
 |
 |
(copyright Peter Brown 1981)
![]()
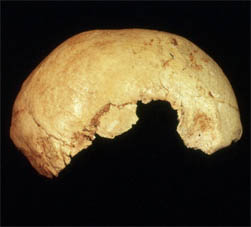
While discovered more than 20 years ago Lake Mungo 3 has not been described in detail. Bowler and Thorne (1976) limit their morphological description to a brief discussion of preservation and the factors which indicate age at death and sex. While photographs of the burial in situ suggest reasonable preservation this was not the situation post excavation. The cranial vault is without most of its left side and base, with the facial skeleton also incomplete. However, the mandible is reasonably complete, apart from some damage to the right condyle. All of the major limb bones, except the right ulna, have damaged or missing articular surfaces and the smaller and more delicate bones of the hands, feet, ribs and vertebral column are particularly poorly preserved. The extent of post depositional bone loss limits the amount of information that can be gained from the skeleton. For example, there is insufficient information available to be absolutely certain of the sex of Lake Mungo 3 (Brown 1987). Diagnostic parts of the pelvic girdle and cranium are not present. It is clear, however, that Lake Mungo 3 was an older adult. There is acute tooth wear and pulp exposure, as well as osteoarthritis in the lumbar vertebrae, right elbow and wrist, with some eburnation (Webb 1989). The right ulna has a maximum length of 297 mm. Using stature estimation formulae from a population with similar relative limb lengths (Lundy, 1983) provides a stature of approximately 170 cm. This would be extremely tall for an Aboriginal woman, from south-eastern Australia, at European contact (Birdsell 1993), but there is evidence of increased stature and skeletal robusticity in the Pleistocene (Brown 2009).
The Mungo 3 cranium is missing the cranial base anterior of opisthion, most of the left temporal, right temporal, and about one third of the right side of the vault. The left supraorbital region is preserved but not glabella. By Australian Aboriginal standards supraorbital development is feminine (supercilliary ridges and zygomatic trigones) and there is no evidence of a median frontal ridge or prebregmatic eminence. The frontal is filled out and not receding. The frontal bone is reasonably thick mid-frontal (11.1 mm) but thins slightly towards bregma (8.6 mm). Most of the cranial sutures are completely fused endocranially, although still visible, which is consistent with the tooth wear and osteoarthritis in suggesting advanced age. The parietals are long and without pronounced antero-posterior curvature. There is a marked depression in the midline of the parietal 35 mm posterior to bregma extending over about 40 mm. Maximum bi parietal breadth is located inferiorly, but this is a reasonably common location in Aboriginal crania. Areas of nuchal muscle insertion are moderate, however, the occipital bone is thickened, particularly at the external occipital protuberance. Viewed laterally, there is slight bunning of the occipital.
More of the left side of the face is preserved than the right. Articulation of the zygomatic with the frontal suggests a fairly rectangular orbit, with only moderate development of the malar tuberosity on the zygomatic. There are a number of maxillary fragments, with some alveolar resorption (incisor) due to tooth loss. Tooth wear is particularly marked in the mandible, with both canines probably lost due to acute wear and there is apical abscess development, and pulp exposure, associated with the first and second molar teeth. The mandibular condyles do not have any evidence of arthritic modification. There is slight gonial eversion in the mandible but the masseteric fossae are not rugose or deep. Anteriorly the mandible has a reasonably prominent mental trigone, but overall chin projection is neutral.
Lake Mungo 3 was further discussed by Thorne (1977) where in a diagram comparing the morphological extremes of late Holocene and "fossil" Australian crania, the latter group containing mid-Holocene and terminal Pleistocene skeletons, LM3 was placed in the middle of the modern female range and outside the male range . Thorne remarks that if it "were it not for femoral and pelvic evidence, one could be tempted to diagnose Lake Mungo III as female" (1977:190). It is unfortunate that Thorne presents no details of this evidence. The division of the "fossil sample" into two contrasting groups was based on the multivariate study of Thorne and Wilson (1977). Lake Mungo 3 had not been available for this analysis but was included in Thorne's descriptive figure to indicate "its probable position" in relation to his recent and fossil samples (1977:190). While details of the gracile, morphologically delicate or modern features of LM3 are not provided Thorne (1977: 191) states that "all of the Lake Mungo individual...possess very thin cranial vault bones" this contrasts with both late Holocene and other late Pleistocene crania such as Kow Swamp. The actual relationship of LM3 to any Australian skeleton, or broadly defined modern humans, has not been tested in Thorne's subsequent publications mentioning this skeleton. More recent comparison of the Australian Pleistocene sample, including LM3, did not support Thorne's division of the sample into two biologically distinct groups (Brown 1987). Thorne et al. (1999), ignoring earlier work to the contrary (Thorne and Wilson 1977; Brown 1987), have again allocated Lake Mungo 3 to a distinctive gracile and modern Pleistocene group. Thorne (1980, 1984, 1989) has argued that this gracile group was the result of a migration from the East Asian region, while the robust group were the descendants of Javan Homo erectus.
While Lake Mungo 3 was available for study by Alan Thorne for several decades, unfortunately only limited detailed information was ever published (Bowler et al. 1970; Thorne 1971, 1976, 1977; Brown 1987a). The reasons behind the inadequate publication of this important early Australian, as well as LM1 and the Kow Swamp remains, are something of a mystery. Without detailed description, as well as radiographs and ct scans, it is difficult to assess the value of the published references to these materials.
![]()
Data file, access and casts
For several decades Lake Mungo 3 was housed at the Australian National University under the care of the late Alan Thorne. Recent newspaper reports (June 1999) indicate that Aboriginal Communities in south-western NSW would like LM3 and the other Willandra Lakes skeletons returned for reburial. These requests intensified after the publication of photographs of LM3 in Australian newspapers in 2001, and the claim that Adcock et al (2001) mtDNA results removed LM3 as an ancestor of Aboriginal people living in the Willandra Lakes region today. Casts of the LM3 skeleton are not available. At the end of this page is a list of metric data for LM3.
![]()
References
Adcock, G., Dennis, E., Easteal, S., Huttley, G., Jermin, L., Peacock, W. & Thorne, A. 2001a. Mitochondrial DNA sequences in ancient Australians: Implications for modern human origins. Proceedings National Academy of Science, 98(2): 537-542.
Adcock, G., Dennis, E., Easteal, S., Huttley, G., Jermin, L., Peacock, W. & Thorne, A. 2001b. Response. Science 292: 1656.
Bowler, J. M., and Thorne, A. G. (1976). Human remains from Lake Mungo: Discovery and excavation of Lake Mungo III. In (R. L. Kirk and A. G. Thorne, Eds.)The Origin of the Australians, pp. 127-138. Canberra: Australian Institute of Aboriginal Studies.
Bowler, J.M. and Magee, J.( 2000) Redating Australia's oldest human remains: A sceptics view. Journal of Human Evolution 38: 719-726.
Bowler, J.M., Johnston, H., Olley, J.M., Prescott, J.R., Roberts, R.G., Shawcross, W. and Spooner, N.A. (2003) New ages for human occupation and climatic change at Lake Mungo, Australia. Nature 421: 837-840. (pdf copy)
Birdsell, J.B. 1993. Microevolutionary patterns in Aboriginal Australia: a gradient analysis of clines. Oxford UP, New York.
Brown, P. (1987). Pleistocene homogeneity and Holocene size reduction: the Australian human skeletal evidence. Archaeol. Oceania 22, 41-71.
Brown, P. (1996). Australian Palaeoanthropology. In F. Spencer (ed.) History of Physical Anthropology: An Encyclopedia. New York: Garland Publishing, pp. 138-145.
Brown, P.( 2000a) Pleistocene variation and the sex of Lake Mungo 3. Journal of Human Evolution 38: 743-750. (pdf copy)
Brown, P.( 2000b) The first Australians. Australasian Science 21 (4): 28-31. (pdf copy)
Brown, P. and Gillespie, R. (2000) On the age and sex of Lake Mungo 3 Australasian Science 21 (5): 26-27.
Brown, P. (2009). Nacurrie 1: the mark of ancient Java or a caring mother's hands in terminal Pleistocene Australia. Journal of Human Evolution 59: 168-187.
Caddie, D. A., Hunter, D. S., Pomery, P. J. and Hall, H. J. (1987). The ageing chemist - can electron spin resonance (ESR) help. In (W. Ambrose and J. Mummery, Eds.) Archaeometery: Further Australasian Studies, pp. 167-176. Canberra: Department of Prehistory, Australian National University.
Cooper, A, Rambaut, A, Macaulay, V, Willerslev, E, Hansen, J & Stringer, C. 2001. Human origins and ancient human DNA. Science 292: 1655-1656.
Gillespie, R. and Roberts, R.G. ( 2000) On the reliability of age estimates for human remains at Lake Mungo. Journal of Human Evolution 38: 727-732.
Gillespie, R. (2002) Dating the first Australians. Radiocarbon 44 (2): 455-472. (pdf copy)
Grün, R. and Schwartz, H. (1987). Some remarks on 'ESR dating of bones'. Ancient TL 5, 1-9.
Nolch, G. 2001. Mungo Man's DNA shakes the Homo family tree. Australasian Science 22 (2): 29-31. (pdf copy)
Smith CI, Chamberlain AT, Rile MS, Stringer C, and Collins MJ (2003) The thermal history of human fossils and the likelihood of successful DNA amplification. Journal of Human Evolution 45:203-217.
Stone T, and Cupper ML (2003) Last Glacial Maximum ages for robust humans at Kow Swamp, southern Australia. Journal of Human Evolution 45:1-13.
Thomas, M., Gilbert, P., Willerslev, E., Hansen, A.J., Barnes, I., Rudbeck, L., Lynnerup, N. and Cooper, A. (2003) Distribution patterns of postmortem damage in human mitochondrial DNA. American Journal of Human Genetics 72: 32-47. (pdf copy)Thorne, A. G. (1976). Introduction. In R.L. Kirk and A.G. Thorne (eds.) The Origin of the Australians. Canberra: AIAS, 1-10.
Thorne, A. G. (1977). Separation or reconcilliation? Biological clues to the development of Australian society. In (J. Allen, J. Golson and R. Jones, Eds.) Sunda and Sahul, pp. 187-204. London: Academic Press.
Thorne, A. G. (1980). The longest link: human evolution in Southeast Asia and the settlement of Australia. In (J. J. Fox, R. C. Garnaut, P. F. McCawley and J. A. C Mackie, Eds.) Indonesia: Australian Perspectives, pp. 35-44. Canberra: Research School of Pacific Studies, A.N.U.
Thorne, A. G. (1984). Australia's human origins - how many sources? Am. J. Phys. Anthropol. 63, 227.
Thorne, A. G. (1989). The emergence of the Pacific peoples. In (L.H. Schmitt, L. Freedman and N.W. Bruce Eds.) The Growing Scope of Human Biology, pp. 103 111. Perth: Australasian Society for Human Biology.
Thorne, A. G. and Wilson, S. R. (1977). Pleistocene and recent Australians: a multivariate comparison. J. hum. Evol. 6, 393-402.
Thorne, A., Grün, R., Mortimer, G., Spooner, N. A., Simpson, J. J., McCulloch, M., Taylor, L. and Curnoe, D. 1999. Australia's oldest human remains: age of the Lake Mungo 3 skeleton. J. hum. Evol. 36, 591-612.
Webb, S.G. (1989). The Willandra Lakes Hominids. Canberra: Department of Prehistory, Australian National University.
![]()
| glabella-opisthocranion | (191) |
| max. bi-parietal breadth | (132) |
| bregma-lambda | 114 |
| parietal subtense height | 18 |
| bregma-subtense point | 65 |
| bregma-asterion | 135 |
| bregma-opisthion | 153 |
| lambda-opisthion | 97 |
| occipital subtense height | 30 |
| lambda-subtense point | 44 |
| asterion-opisthion | 71 |
| bi-asterion | (113) |
| lambda-asterion | 79 |
| thickness at mid-frontal | 11.1 |
| thickness at bregma | 8.6 |
| thickness at vertex | 8.0 |
| thickness at parietal boss | 6.6 |
| thickness at asterion | 8.0 |
| thickness at lambda | 8.7 |
| th. at external occip. protuberance | 15.5 |
| mandibular right I2 b-l breadth | 7.0 |
| mandibular right PM1 b-l breadth | 9.3 |
| mandibular right PM2 b-l breadth | 9.7 |
| mandibular left M3 b-l breadth | 11.8 |
| symphyseal height | 35 |
| symphyseal thickness | 16 |
| bi-gonial breadth | 106 |
| bi-condylar breadth | (129) |
| ramus minimum breadth | 42 |
| mandible length | 111 |
![]()